8 دقیقه
تحقیقات ژنتیکی تازه نشان میدهد که گامهای آغازین بهسمت سلولهای پیچیده و دارای هسته — جدّ اجداد همه موجودات از آمیباها تا انسانها — تقریباً سه میلیارد سال پیش برداشته شده است. این بازتاریخگذاری مبدا پیچیدگی یوکاریوتی را تا حدود یک میلیارد سال به عقب میبرد و نشان میدهد که یک تجمع تدریجی از نوآوریهای مولکولی و ساختمانی مدتها پیش از اکسیژندار شدن گستردهٔ زمین رخ داده است.
بازاندیشی درباره زمان آغاز پیچیدگی
زندگی روی زمین معمولاً در دو گروه کلی دستهبندی میشود: پروکاریوتها (باکتریها و آرکئا) و یوکاریوتها (سلولهایی با هسته و اندامکها). پروکاریوتها ابتدا پدید آمدند، تقریباً چهار میلیارد سال پیش، بهصورت سلولهایی جمعوجور و کارآمد با DNA شناور و ساختار داخلی محدود. در مقابل، یوکاریوتها دارای غشاهای داخلی، هسته و اندامکهایی مانند میتوکندری هستند که از ژنومهای بزرگتر، بیان ژنی تنظیمشدهتر و فرآیندهای سلولی پیچیدهتر پشتیبانی میکنند.
سؤال اساسی که در طول دههها بحثبرانگیز بوده این است که جهش به سوی این پیچیدگی دقیقاً چه زمانی و چگونه رخ داده است. یکی از محورهای مناقشه زمانبندی «تسخیر میتوکندری» است — فرایند آندوسیمبیوتیک باستانی که در آن یک باکتری آزادزی به کارخانهٔ تولید انرژی سلول تبدیل شد. آیا میتوکندریها محرک اصلی پیدایش ویژگیهای یوکاریوتی بودند یا پروتو-یوکاریوتها ابتدا بخشی از ساختارهای درونی را ایجاد کردند و سپس میتوکندری را جذب کردند؟
مفهومهای جدید از جمله کشف گروههایی مانند آرکئاهای متعلق به ابرگروه Asgard (مثلاً Lokiarchaeota) نشان میدهد که برخی ویژگیهای مولکولی مرتبط با یوکاریوتها ممکن است در خطهای باستانی آرکئا پیشنهاد شده باشند. این یافتهها با ایدهٔ یک سیر تکاملی تدریجی هماهنگ است که در آن ماژولهای مولکولی مختلف بهتدریج یکپارچه شده و زمینه را برای ساختارهای پیچیدهتر فراهم کردند.
چگونه ساعت مولکولی جدول زمانی را بازنویسی میکند
برای روشن شدن این موضوع، تیمی به رهبری دیرینهشناس زیستی کریستوفر کی از دانشگاه بریستول یک تحلیل گستردهٔ ساعت مولکولی را روی صدها گونه اجرا کردند. ساعتهای مولکولی با مقایسهٔ توالیهای DNA یا پروتئین و بهکارگیری نرخهای جهشی که با شواهد فسیلی کالیبره شدهاند، زمانهای جدایی و ظهور ویژگیها را تخمین میزنند. این پروژه میانرشتهای، با تلفیق فیلوژنتیک، دیرینهشناسی و زیستشناسی مولکولی، تلاش کرد تا ظهور خانوادههای ژنی ویژه را روی یک جدول زمانی مطلق قرار دهد.
ازجمله نکات روششناختی این مطالعه، استفاده از مجموعهٔ گستردهای از خانوادههای ژنی بهجای اتکا به تنها چند ژن نشانگر است. پژوهشگران هزاران توالی پروتئینی را مورد بررسی قرار دادند، تعاملات پروتئینی را تحلیل کردند و خانوادههای ژنی را بر اساس عملکرد و شبکهٔ تعاملات گروهبندی نمودند. سپس با استفاده از مدلهای ساعت مولکولی رها (relaxed clock) و نقاط کالیبراسیون فسیلی مختلف، زمان تقریبی پیدایش هر ماژول مولکولی را برآورد کردند.
در این فرآیند، مواجهه با چالشهایی مانند انتقال افقی ژن (HGT)، تغییرپذیری نرخ تکاملی بین شاخهها و اثرات جذبِ بلند-شاخه (long-branch attraction) نیازمند استفاده از روشهای پیچیدهٔ آماری و اعتبارسنجی متقابل بود. بهطور کلی، استفاده از نمونهبرداری تاکسونومیک گسترده و کالیبراسیونهای زمینشناسی متنوع به پویایی مدل و کاهش سوگیریها کمک کرد.
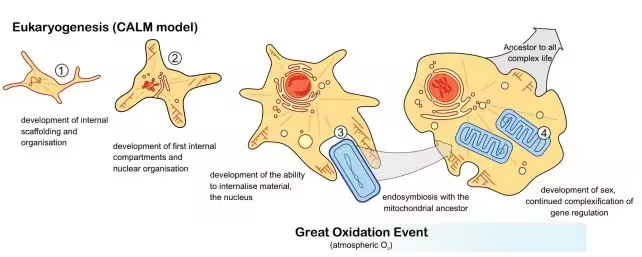
خط زمانی تیم پژوهشی برای تکامل یوکاریوتها
مدل توسعهیافته توسط گروه با عنوان CALM — مخفف Complex Archaeon, Late Mitochondrion — طراحی شد تا مشخص کند کدام ویژگیهای یوکاریوتی در چه زمانی ظاهر شدهاند. بر خلاف مطالعات محدودتر که تنها چند ژن شاخص را مدنظر قرار میدهند، CALM صدها خانوادهٔ ژنی مرتبط با ساختارها و فرآیندهای کلیدی یوکاریوتی را دنبال کرد تا تصویر زمانی جامعتری ارائه دهد.
یافتههای کلیدی: زیرساختهای اولیه، میتوکندری دیرهنگام
نتایج مطالعه قابل توجه است. الگوهای مولکولی مربوط به پروتئینهایی که در شکلگیری سیتواسکلتون نقش دارند — از جمله آکتین و توبولین — همراه با شواهدی از ساختارهای ابتدایی سیتواسکلتال و شکلگیری یک پروتو-هسته (incipient protonucleus) تقریباً بین ۲٫۹ تا ۳٫۰ میلیارد سال پیش ظاهر میشوند. این یافته نشان میدهد که زیرساختهای سازمانی داخلی سلولها، مانند شبکههای فیلامنتی و سیستمهای جابهجایی درونسلولی، بسیار زودتر از انتظار قدیمی به وجود آمدهاند.
موجهای بعدی نوآوری شامل پیدایش غشاهای داخلی پیچیدهتر، اجزایی مرتبط با دستگاه گلژی و گسترش سیستمهای بیان ژنی — از جمله تکثیر انواع RNA پلیمرازها و عوامل رونویسی متنوع — بوده است. این توالی پیدایش، تصویر یک ساختارسازی مرحلهای را نشان میدهد که به تدریج قابلیتهای مدیریت ژنومی و پیچیدگی عملکردی سلول را افزایش داده است.
در تقابل با این روند ابتدایی، شاخهای که منجر به میتوکندریها شد، در زمانی بهطور معنیداری دیرتر — تقریباً ۲٫۲ میلیارد سال پیش — تاریخگذاری میشود. این زمانبندی تقریباً همزمان با رویداد بزرگ اکسیژناسیون (Great Oxidation Event) است، دورهای که در آن سطح اکسیژن جوی بهطور چشمگیری افزایش یافت. این همزمانی نشان میدهد که بسیاری از نوآوریهای یوکاریوتی در شرایط کماکسیژن آغاز شده بودند، ولی ورود میتوکندری و تغییرات ردوکس اتمسفر احتمالاً محرک افزایش سریع تنوع و پیچیدگی پس از آن بوده است.
چرا این یافتهها برای زیستشناسی تکاملی اهمیت دارد
اگر صفات پایهای یوکاریوتها میلیاردها سال پیش از ورود میتوکندریها ظاهر شده باشند، نتیجهٔ مهم این است که تکامل یوکاریوتی یک توالی طولانی و پلهای از نوآوریها بوده است، نه یک جهش واحد و انفجاری. سیستمهای سادهٔ سیتواسکلت و غشاهای جداساز میتوانستند مزایای سازمانی ملموسی ایجاد کنند — مدیریت مؤثرتر ژنوم، هدایت و انتقال درونسلولی بهتر و تنظیم فضایی فرآیندهای زیستی — حتی در غیاب سطوح بالای اکسیژن.
با بالا رفتن سطوح اکسیژن و ورود میتوکندریها، سلولها قادر به بهرهبرداری از متابولیسمهای انرژیزاتر (بهرهوری بیشتر تولید ATP) شدند. این تغییر انرژیِ قابل استفاده، امکان توسعهٔ فرآیندهای پرمصرف انرژی را فراهم کرد؛ در نتیجه ژنومها میتوانستند بزرگتر شوند، شبکههای پروتئینی پیچیدهتر شکل بگیرند و سلولها نقشهای زیستی متنوعتری را در اکوسیستمها بر عهده گیرند.
این دیدگاه همچنین بر بحثهای نظری دربارهٔ هزینه-فایدهٔ تکامل ساختارهای درونی سلولی تأکید میکند: شکلگیری غشاها و ماژولهای ساختاری میتواند بدون مزایای انرژی مستقیم نیز بهعنوان سرمایهگذاری سازمانی اولیه عمل کند که بعدها با افزایش انرژی جان دوباره میگیرد.
عمق روششناختی و کار میانرشتهای
یکی از تمایزهای این مطالعه وضوح در سطح خانوادههای ژنی همراه با پیوند به زمانهای مطلق است. با تحلیل شبکههای تعامل پروتئینی، گروهبندی عملکردی خانوادههای ژنی و استفاده از نقاط کالیبراسیون فسیلی متعدد (از جمله فسیلهای میکروبی و سنگوارههای مولکولی مانند نشانگرهای استرانیها)، نویسندگان نه تنها یک درخت جدایی را بازسازی کردند بلکه توالی زمانی ظهور سامانههای سلولی را بازسازی نمودند.
این کار مستلزم همکاری نزدیک میان دیرینهشناسان برای تأمین محدودیتهای سنی، فیلوژنتیستها برای ساختن درختهای قوی و زیستشناسان مولکولی برای تفسیر عملکرد ژنها بود. همچنین تحلیل حساسیت نسبت به فرضیات کالیبراسیون، ارزیابی سناریوهای مختلف انتقال افقی ژن و بررسی مدلهای سرعت تکاملی متفاوت، جزء لاینفک استحکام نتایج بود.
بهعلاوه، چنین مطالعهای نشان میدهد که چگونه تلفیق دادههای ژنومی با شواهد زمینشناسی و ژئوشیمیایی میتواند به پرسشهای دقیقتری پاسخ دهد: کدام ماژولهای مولکولی نخستین شکل گرفتند؟ کدام ماژولها بعداً بهکار گرفته شدند؟ و چگونه تغییرات محیطی مانند افزایش اکسیژن یا تغییر در دسترسی به مواد مغذی، مسیرهای تکاملی را جهتدادند؟
پیامدها و مسیرهای پژوهشی آینده
این نتایج دید ما از تاریخ عمیق حیات را بازتنظیم میکنند. آنها نشان میدهند که سازماندهی سلولی پیچیده میتواند بهتدریج و در شرایط کماکسیژن شکل بگیرد و تنها پس از افزایش انرژی قابلاستفاده سریعتر گسترش یابد. این نکته برای زیستشناسی فرازمینی و جستجوی حیات در سیارات دیگر نیز اهمیت دارد: پیچیدگی زیستی ممکن است در مراحل متعدد شکل گیرد و لزوماً همزمان با اکسیژندار شدن سیاره نباشد.
تحقیقات بعدی میتوانند تخمینهای زمانی را با دقت بالاتری اصلاح کنند، مدل CALM را در برابر ژنومهای بیشتر و تنوع تاکسونومیک گستردهتر آزمون کنند و نقش دقیق متغیرهای محیطی — مانند در دسترس بودن مواد مغذی، وضعیتهای ردوکس محلی و ساختارهای جمعیتی میکروبی — را بررسی نمایند. همچنین، رکوردهای سنگی بسیار کهن، نشانگرهای مولکولی (biomarkers) جدید و ژنومهای باکیفیت از شاخههای میکروبی متنوع، کلیدهای اصلی برای کشف جزئیات بیشتر خواهند بود.
از منظر فنی، بهبود مدلهای ساعت مولکولی (از جمله مدلهای مبتنی بر Bayesian و مدلهای با نرخهای متغیر بین شاخهای)، افزایش نمونهبرداری تاکسونومی و ترکیب شواهد مولکولی با دادههای پترولوژیکی و ژئوشیمیایی، مسیرهایی است که میتواند اعتبار و دقت نتایج را افزایش دهد.
دیدگاه کارشناسی
"این نتایج تصویری از نوآوری تدریجی را ترسیم میکنند تا تحولی آنی،" میگوید دکتر Elaine Moreno، زیستشناس تکاملی که در این مطالعه دخالتی نداشته است. "ظهور اجزای سیتواسکلت و سامانههای غشایی در این دوران بسیار دیرینه نشان میدهد که انتخاب طبیعی مدتها پیش از ورود میتوکندریها در معماری داخلی سلولها دستبهآزمایش میزد. وقتی اکسیژندار شدن و آندوسیمبیوز میتوکندری رخ داد، آن سامانههای پیشساز قادر بودند به سرعت رشد و مقیاسپذیری پیدا کنند."
او میافزاید: "از منظر کاربردی، این کار ارزش ترکیب شواهد فسیلی، درختان ژنی مبتنی بر عملکرد و نمونهبرداری گستردهٔ تاکسونومیک را برجسته میکند. این روش الگویی قدرتمند برای مطالعهٔ گذارهای عمیق تکاملی فراهم میآورد و میتواند در بررسی سایر رویدادهای بنیادی زیستی نیز بهکار رود."
چه مواردی را دنبال کنیم
- گسترش نمونهبرداری ژنومی از آرکئا و یوکاریوتهای شاخهدار عمیق (deep-branching eukaryotes) برای آزمون فراگیری نشانگرهای اولیهٔ یوکاریوتی و تأیید توالیهای پیشنهادی ظهور ماژولها.
- مطالعات ژئوشیمیایی و پترولوژیک برای دقیقتر کردن زمانبندی و تعیین تغییرپذیری منطقهای افزایش اکسیژن در زمین اولیه؛ از جمله تحلیل ایزوتوپها و نشانگرهای زیستی که ارتقای مرحلهای اکسیداسیون را منعکس میکنند.
- کارهای تجربی بازسازی پروتئینهای اجدادی (ancestral protein reconstruction) برای ارزیابی عملکرد اجزای سیتواسکلت اولیه در محیطهای کماکسیژن و بررسی خواص سینتیکی و پایداری آنها.
- تحقیقات در زیستشناسی سیستمی و مدلسازی انرژی (energetic budget modeling) تا روشن شود چگونه افزایش منابع انرژی پس از ورود میتوکندری به گسترش شبکههای مولکولی و رشد ژنومی کمک کرد.
ادغام این رویکردها با دادههای زمینشناسی و مدلهای اکوسیستمی میتواند تصویر بسیار دقیقی از سیر تکاملی پیچیدگی سلولی ارائه دهد و پرسشهای جدیدی در زمینهٔ منشاء یوکاریوتها، زمانبندی ظهور اندامکها و شرایط محیطی مؤثر بر این تحولات مطرح نماید.





.webp "نخستین نشانه جدی از قمر فراخورشیدی عظیم دوردست")






نظر بگذارید
نظرات (7)
اوه… عجب! این یعنی احتمالا روی سیارات بیاکسیژن هم ممکنه یوکاریوتنماها شکل بگیرن؟ 😮
شاید کمی اغراق شده، بازسازی ها و کالیبراسیونها حساسن... اما ایدهٔ مرحلهای منطقیه
نقطه نظر خوبیه، ترکیب فسیلی و ژنومی قوی، ولی ساعت مولکولی هنوز میتونه سوگیری داشته باشه
تو آزمایشم دیدم که پروتو-سیتواسکلتون ها تو شرایط کم اکسیژن هم کار میکردن، جالب اما سخت اثباتشه
این HGT رو چطور تصحیح کردن؟ اگر انتقال افقی زیاد باشه، زمانبندی به هم نمیخوره؟
منطقیشه، مرحله ای بوده نه جهش ناگهانی. اما سندها رو ببینیم، خیلی چیزا هنوز مبهمه
واااای، سه میلیارد سال؟! یعنی سلولها کلی قبل از اکسیژن پیچیده شده بودن، فکرشو نمیکردم...